Annona cherimola, la
chirimoya o
chirimoyo (<
quechua chiri, "frío, fría",
muya, "semillas", puesto que germina a elevadas altitudes) es una
especie de
árbol perteneciente a la familia de las
annonáceas, nativa del Perú y Ecuador,
América del Sur. Ya era cultivado en los
Andes del
Perú desde el año
200 por la
cultura Mochica.
[1
Es un
árbol frutal originario de los valles interandinos del
Perú y
Ecuador, de áreas comprendidas entre los 1.500 y 2.200
msnm de altitud. Se introdujo en el sur de
España antes de
1751,
[2] en la zona de
Motril,
Almuñécar y parte
Málaga, de donde probablemente se llevó a
Italia e lsla de
Madeira (
Portugal). En la zona mediterránea también se encuentra bien adaptado en
Israel,
Egipto,
Grecia y
Argelia.
Características

Fruto de la
Annona cherimola.
El chirimoyo es un árbol de crecimiento lento, que puede adquirir en su madurez una altura de 7 a 8 m, y exuberante follaje; de porte erguido y a veces ramificado irregularmente. El
tallo es cilíndrico, de corteza gruesa. Posee un sistema radicular muy superficial y ramificado, originando dos o tres pisos a diferentes alturas, pero poco profundos.
Las
hojas son simples, enteras, de disposición alterna y de forma ovalada u ovada-lanceolada. Las yemas son compuestas y pueden originar brotes mixtos (vegetativos y florales).
Las flores, de seis
pétalos amarillentos jaspeados de púrpura,
hermafroditas, son muy aromáticas, poco llamativas, solitarias o en ramilletes de dos o tres, sobre un corto e inclinado
pedúnculo inserto en las axilas de las hojas. El cáliz consta de 3
sépalos de color verde oscuro, pequeños y de forma triangular. La corola está formada por seis pétalos dispuestos en dos verticilos; los tres pétalos exteriores bien desarrollados son carnosos, miden de 2,5 a 4 cm de longitud y la parte superior tiene forma aquillada o triangular; los tres pétalos internos son rudimentarios, en forma de escama, ovalados o triangulares. La parte masculina de la flor consta de numerosos
estambres (150-200), dispuestos helicoidalmente muy juntos sobre un receptáculo, formando una masa compacta y blanca oprimida por los pétalos. La parte femenina posee también elevado de número de carpelos (de 100 a 200), con un solo óvulo, dispuestos en espiral, formando un cono compacto en cuyos extremos se encuentran los estilos y estigmas. Al fecundarse los óvulos se desarrolla un fruto compuesto (sincarpo), como consecuencia de la fusión de los
carpelos, alrededor de un receptáculo carnoso de forma alargada y cónica. Cuando la polinización es inadecuada y sólo se fecundan algunos óvulos de manera irregular, los frutos que se forman son asimétricos y deformes. La piel es fina y delicada, la superficie del fruto presenta marcas en forma de U que se corresponden con la zona de unión de los carpelos, pudiendo ser lisa o con pequeñas protuberancias. El peso puede oscilar entre 200 y 800 g. El color del fruto, según la variedad, puede ir de verde claro a verde oscuro. La pulpa es blanca, cremosa y moderadamente jugosa, con numerosas semillas de color desde marrón oscuro a negro; el sabor es subácido y delicado.
Como en todas las especies de esta familia, éste no es un fruto simple sino un agregado de frutos adheridos sobre un sólo receptáculo, producto de las pequeñas
flores que se fecundan por separado. Así, la chirimoya perfecta es acorazonada y sólo se consigue en condiciones ideales de
polinización, abundante y uniforme, hecha por escarabajos, como la mayoría de las especies pertenecientes al orden
magnoliales, o por la mano humana. Su tamaño es similar al
pomelo. La
cáscara es delgadísima y frágil; su superficie verde oscura, casi lisa, lleva como una red de sombras que demarca los límites de cada frutilla. El interior de la fruta, de
color blanco, posee una textura carnosa, blanda y de
sabor dulce, a veces descrita como una mezcla entre la
piña, el
mango y la
fresa.
Valor nutritivo
Su valor
nutritivo se explica por el elevado contenido de
azúcares, que llega hasta el 20%, y el de
proteínas, superior al de muchas otras frutas, que alcanza el 2%. Tiene mucha
vitamina B1,
B2,
calcio (25-32
mg),
hierro (0.5 mg) y
fósforo (37 mg)
Usos
Se dice
[cita requerida], pero aún no está probado, que la chirimoya posee un componente hipotensor que activa la
enzima óxido nítrico sintasa encargada de sintetizar o fabricar
óxido nítrico (N2O5) — un vasodilatador responsable de la erección peneana de forma natural y fisiológica — similar a los efectos provocados por la conocida
viagra.
[cita requerida]
Cultivo
El árbol requiere de climas secos donde no llueva mucho y donde la
temperatura no presente extremos de calor ni de frío. En el trópico busca para crecer alturas entre 1000 y 2000
msnm y es sensible a los ataques de
hongos y otras plagas.
Los emplazamientos sin grandes fluctuaciones de temperatura y humedad son los más propicios para este cultivo. Las condiciones ideales para obtener un buen cuajado de frutos son temperaturas entre 25 y 28 °C y humedades relativas entre 60-70 % durante la época de floración. La temperatura media que se acepta como límite para el cultivo de esta especie es la de 13 °C, en los meses más fríos. Por debajo de esta temperatura existe mayor dificultad para obtener frutos comestibles. Es un árbol muy susceptible a las heladas y a temperaturas por debajo de -2 °C, se pueden ocasionar daños tanto en la madera como en las hojas y frutos, irreparables si estas temperaturas permanecen durante algunas horas. En época de floración las temperaturas superiores a 30 °C, junto con bajas humedades relativas, afectan negativamente a la fecundación al provocar pérdida de receptividad estigmática.
Los vientos fuertes también son perjudiciales para el chirimoyo ya que sus ramas más jóvenes pueden doblarse por acción del mismo, provocando una mala formación del árbol. También el viento puede tener un efecto negativo sobre los frutos en su etapa de maduración en el árbol, ya que la piel puede verse dañada debido a los roces que se producen con la ramas.
El chirimoyo, igual que las demás especies del género
Annona que se cultivan, se adapta a diversos tipos de suelos, como pueden ser arenosos, limo-arenosos, o arcillosos, e incluso se ha visto que crecen bien en suelos pedregosos, con la única exigencia de que éstos tengan un buen drenaje, puesto que no soportan los encharcamientos.
El pH del suelo más adecuado, de acuerdo a la literatura, sería el comprendido entre 6,0 y 7,5.
Fertilización
Es importante dar una aplicación alta de estiércol (5
t/
ha) bien descompuesto una vez por año. Tomando en cuenta los análisis de suelo realizados, se determinaran las dosis de los fertilizantes. Así se realizan aplicaciones de fertilizantes a base de N-P-K. A medida que la planta se va desarrollando incrementar la dosis de nitrógeno. Se debe tener en cuenta la frecuente carencia de potasio en este cultivo, con la aparición de márgenes foliares con decoloraciones que evolucionan a necrosis. Las necesidades de calcio y magnesio son importantes sobre todo en la última fase de crecimiento del fruto, previo a la maduración
Un cultivo de 14 tm/ha extrae:
- Nitrógeno (N) . . .95 kg/ha
- Fósforo (P) . . . .4,6 kg/ha (11,3 kg/ha de P2O5)
- Potasio (K) . . . .38 kg/ha (45 kg/ha de K2O)
- Calcio (Ca) . . . .9 kg/ha
- Magnesio (Mg) . .7,5 kg/ha
Poda
Es una planta muy competitiva por la luz para la producción, por lo que es conveniente la poda de formación, dejando una copa equilibrada de formas bastante libres abriendo el centro a la luz e intentando, además, facilitar la polinización artificial y la recolección. Tras la recolección se da una poda de mantenimiento la cual consiste en eliminar ramas torcidas, ramas verticales y chupones, dejándole una máxima expansión.
Poda de formación
Se lleva a cabo en los tres primeros años, con el fin de obtener la adecuada estructura del árbol que le permita soportar, posteriormente, una buena carga. En general se realiza la poda en vaso con 3 a 5 ramas principales, formadas a 1 m de altura, aproximadamente. Con esta poda de vaso bajo se disminuyen los problemas con el viento, se facilita la recolección y se aumentan los rendimientos de la polinización manual, donde ésta sea necesaria. La estructura del árbol deberá estar formada al tercer año, altura en la que el árbol comienza a producir los primeros frutos.
Poda de mantenimiento (a finales de invierno)
Consiste en general, en eliminar primero todo el material seco y aquel que esté creciendo hacia el interior y que no tenga grandes posibilidades de recibir abundante luz. Como las flores aparecen preferentemente en las ramas del año anterior, se deberá tener cuidado de no eliminar este material que es el más productivo. Se sabe que los frutos de mayor peso y calidad son los producidos en las ramas con dos o más años (aunque en menor número) las cuales fructificarán si eliminamos parte de las ramas del año anterior.
Poda de fructificación
Cada año, a la salida del invierno y antes del inicio de la nueva brotación, se procede a un aclareo de ramas fructíferas y despunte de ramas principales. Durante el periodo vegetativo se procede al pinzamiento o eliminación de los chupones. En la actualidad, las plantaciones antiguas tienen una densidad de 150 árboles por hectárea con alturas de 4 a 4,5 m .
Deshierba
Es frecuente el laboreo con vertedera, con cuidado de no dañar las raíces superficiales ya que es muy susceptible a fitóftora. Es muy recomendable mantener el suelo cubierto, preferentemente con plantas leguminosas entre los árboles, que por su aporte de nitrógeno resultan las mejores.
a) Mantener la fila de árboles limpia de malas hierbas mediante la aplicación de herbicidas de preemergencia a través del riego por microaspersión (Goal (Oxifluorfeno) y Simazina en 12 aplicaciones por año). Las hierbas perennes que escapan a este herbicida se eliminan con Glifosato ó Paraquat + Diquat.
b) En las calles se deja la vegetación natural, segada 3 veces al año. Debe tenerse en cuenta que por falta de lluvias, la hierba no crece en verano en las zonas no regadas.
Propagación
Se lleva a cabo mediante injerto sobre patrón de semilla procedente del mismo cultivar que se va a utilizar como variedad. Se recomienda el injerto en bisel o el de púa empleando los chupones, para obtener mayor estabilidad. Cuando las plantas adquieren, como mínimo, de 1,5 a 2 cm de grosor (medido aproximadamente a 25 cm del suelo) se injertan de yema o de púa lateral o terminal, a unos 50 cm del suelo. Una vez que los árboles alcanzan una altura de 1 m sobre el punto del injerto se plantan en el terreno definitivo, recomendándose que se realice mientras el árbol esté sin hojas (finales del invierno - principios de la primavera) y antes de que haya comenzado la brotación.
Fisiopatías y desórdenes físicos
Daño por frío: la exposición de frutos de
Annona a temperaturas inferiores a 8-12 °C (46-54 °F), dependiendo del cultivar y estado de madurez, resulta en daño por frío. Los síntomas incluyen un oscurecimiento y endurecimiento de la cáscara, depresiones, incapacidad de desarrollar buen sabor, y pulpa "harinosa".
Partiduras: en algunos cultivares la partidura de frutos ocurre en estados avanzados de madurez y con tasas altas de producción de etileno. Se ha sugerido que cambios en la turgencia relacionados a la producción de azúcares neutros durante la maduración provoca movimientos de agua desde la cáscara y posiblemente del receptáculo hacia la pulpa. El incremento en el diámetro del receptáculo aumenta el estrés en la pulpa y cáscara, lo que produce la partidura del fruto.
Enfermedades
En general son pocas las plagas que afectan a este cultivo, destacando por su importancia económica la de
Ceratitis capitata (Wied) (mosca mediterránea o de la fruta) que produce graves daños en los frutos (Málaga y Granada). En Canarias las plagas más frecuentes son la de
Planoccocus citri (cochinilla algodonosa) y la de
Saisettia oleae que pueden atacar tanto a ramas, hojas, flores y frutos. Éstas pueden ser controladas impidiendo el acceso al árbol de la hormiga argentina (
Iridomyrmex humillis), que es el transporte de estas cochinillas, mediante bandas con Diazinán o similar. Para los tratamientos directos contra estas plagas de cochinillas se recomienda el aceite de verano. Como enfermedad de la raíz y tronco está la provocada por el hongo
Phythopthora cínnamomi. Esta enfermedad es favorecida por suelos excesivamente compactos, encharcados y de difícil drenaje, por lo que en plan preventivo deben evitarse suelos con estas características, así como partir de material vegetal sano. Los fungicidas recomendados para esta enfermedad son Metalaxil, Fosetil-Al. y ácido fosforoso neutralizado con hidróxido de potasio, o similares.
Distribución
La chirimoya es un
árbol caducifolio de la familia de las
anonáceas, nativo del área
subtropical de los
Andes, que se cultiva por su
fruto.
Esta fruta propia de
Sudamérica se cultiva también en
Europa, concretamente en el
sur, en la
costa de Granada (España), en los
municipios de
Almuñécar,
Jete,
Motril,
Otívar y
Salobreña. En la actualidad, España es el primer productor mundial de chirimoya con 80% del total, con denominación de origen las de la Costa de Granada y Málaga. Está naturalmente adaptada a las condiciones locales, convirtiéndose en un cultivo muy rentable pero que ha alcanzado la saturación de un mercado limitado.
Los principales productores son
Argentina,
Australia,
Bolivia,
Chile,
Colombia,
Ecuador,
España,
Estados Unidos,
Israel,
Perú,
Sudáfrica y
Taiwán.
Dentro de esta familia, las especies que producen frutos comestibles son fundamentalmente del género
Annona, muchas de las cuales se cultivan comercialmente en los trópicos. Las más importantes son:
- A. cherimola Mill. (chirimoya),
- A. diversifolia Saff. (ilama),
- A. muricata L. (guanábana) (En Puerto Rico también es conocida como "Pana de pepita"),
- A. purpurea Moc. & Sessé ex Dunal (soncoya),
- A. reticulata L. (corazón), y
- A. squamosa L. (anón).
Otras especies, pertenecientes a otros géneros con frutos comestibles, son:
- Asimina triloba (L.) Dunal (asimina), especie nativa de zonas templadas de Norteamérica, y
- Rollinia mucosa Baill., [Rollinia deliciosa Saff. (biriba).
Existe también un híbrido entre
A. cherimola y
A. squamosa, denominado «atemoya», producido en Florida por Pj. Westery E. Simmonds, hoy en día objeto de cultivo comercial tanto en Florida como en Australia e Israel.
Comercialización
Si bien en el actual territorio de Ecuador, Perú y Bolivia existen especímenes silvestres, la producción de estos países representa una fracción muy baja respecto del total mundial, en razón de que no existe una industria desarrollada, empleándose los métodos tradicionales de cultivo de muy baja productividad por superficie.
En Republica Dominicana la chirimoya es conocida comunmente como Mamón, es de difícil producción y existen diversas especies de "Chirimoyas" como son conocidas en el pais caribeño.
España es el primer productor de chirimoyas a nivel mundial, con una superficie plantada de unas 3.300
ha (en la costa de
Málaga y
Granada) y 30.000 t, aproximadamente, de producción anual. En segundo lugar figura
Chile, donde la superficie plantada es de 1.316 ha, con una producción anual de 5.500 t .
Hasta la fecha, los únicos abastecedores de Europa han sido Chile y España. Chile inició su exportación en
1978 con tan sólo 422
kg enviados a
Alemania Federal. En años posteriores fueron aumentándolas hasta alcanzar en
1991 la cifra de 73.370 kg distribuidos entre
América del Sur (mayormente Argentina), varios países europeos (entre ellos España),
América del Norte (
Canadá) y
Oriente Medio.
La exportación española es también reducida debido a que la demanda interior absorbe casi toda la producción; según fuentes del SOIVRE, en 1990 tan sólo se exportaron 286.159 kg, de los cuales un 64% fue enviado a Francia y el resto a Alemania,
Reino Unido, Portugal y otros. Los demás países productores comercializan su fruta en el propio país.
El futuro de mercado para esta fruta, tras la incorporación de España a la
U.E., parece prometedor, teniendo en cuenta la repercusión negativa que supone la superproducción de algunas de las especies frutícolas tradicionales difundidas por Europa, que hacen aumentar el interés por frutos poco conocidos. El chirimoyo es un árbol frutal con grandes posibilidades de desarrollo en la franja Mediterránea. Por ello, los países que la integran (España, Italia, Grecia, Portugal, Israel,
Chipre y
Turquía), están actualmente dirigiendo sus esfuerzos hacia una selección varietal y programas de mejora, con el fin de implantar este cultivo de cara a introducir sus frutos en la U.E., hasta hace poco, como se ha dicho, abastecida en escasa medida por Chile y España.
En la actualidad, el chirimoyo presenta una concentración excesiva de la oferta desde primeros de octubre a finales de noviembre, más del 60%, (Calatrava, 1998), debido, fundamentalmente, al cultivo de una sola variedad, Fino de Jete, lo que repercute negativamente en los precios obtenidos por el agricultor. Otros problemas son la corta vida comercial del fruto, la concentración de la oferta en determinados mercados y la calidad variable del producto.
La calidad variable de la chirimoya ofertada en el mercado se debe a:
- Mal manejo de las técnicas de producción (poda, polinización, riego, etc.) lo que se traduce en frutos pequeños con alto índice de semillas.
- La recolección antes de la fecha adecuada baja el sabor del fruto y sube el índice de semillas.
- Manipulación inadecuada de la fruta durante la recogida o el manejo en almacén. Los frutos golpeados tienen una mala presentación. El chirimoyo es especialmente sensible a daños en los bordes de las areolas, cuando los frutos ruedan unos sobre otros.
- El chirimoyo se ablanda muy rápidamente tras la cosecha. Por ello, resulta fundamental un enfriamiento rápido hasta 9-10 °C, sobre todo cuando el fruto se destina a la exportación.
En el caso chileno el problema es peor porque exportan a largas distancias cosechando un fruto muy verde que se deteriora con mal sabor en el mercado y en vez de aumentar la venta, causa rechazo de los consumidores en
Florida y una contracción del mercado.
- Chile lo exporta verde para que llegue maduro, y el rechazo en el consumidor se debe al número de semillas, lo que ya se resolvió en la Universidad Católica de Valparaíso(PUCV,Chile),creando frutos partenocárpicos(sin semillas).
Otros usos
Además de su uso en helados, los productores bolivianos han empezado a incursionar en la comercialización de la pulpa de la fruta en frascos que contienen sólo la pulpa y permiten preservar su contenido durante varios meses. También venden helados, licor de chirimoya, dulces y tortas, entre otros.
[1]
Referencias
- ↑ Larco Hoyle, Rafael (2001). Los Mochicas. Lima: Museo Arqueológico Rafael Larco Herrera. ISBN 9972-9341-0-1.
- ↑ La Vanguardia, Barcelona, 19 de junio de 1954, pp.5
Bibliografía
- Mark A. Mossler & O. Norman Nesheim. Oct 2002, Florida Crop/Pest Management Profile: Atemoya and Sugar Apple.
- Morton, J. 1987. Cherimoya. p. 65–69. In: Fruits of warm climates. Julia F. Morton, Miami, FL.
- Farré, J.M., J.M. Hermoso y M.A. González, 1976: Ensayos sobre polinización, cuajado y crecimiento del fruto en chirimoya. 1.- An. INIA/Ser.:Prod.veg. Nº 6:63-92.
- Gardiazábal l., E y G, Rosenberg M., 1993: El cultivo del chirimoyo,-Ed. Univ, de Valparaiso. Univ. Católica de Valparaiso. Chile.: 145p.
- Gil-Albert, F, 1986: La ecología del árbol frutal. - Secr. General Técnica. MinisterioAgricultura, Pesca y Alimentación., Serie Técnica. Madrid: 278 p.
- Guirado. E., 1991: Polinización artificial del chirimoyo. Gab. Técn. Caja Rural de Granada, 15 p. Ibar, L,, 1979: El Chirimoyo.- En: Aguacate, Chirimoyo, Mango, Papaya. Edit. Aedos. Barcelona: 121-144.
- Roseli, P 1995: Estudio sobre la Biologia Floral del chirimoyo (Annona cherímola Mill.) en la isla de Tenerife. Tesis Doctor-al. Univ. La Laguna (Tenerife), 195 p.
- Roseli, P.and V. Galán, 1996: Notes on rhythms observed in the duration of flower anthesis throughout flowering in cherimoya on the island of Tenerife.- Fruits, 50 (3): 233-237.
- Schroeden C.A., 1942: Hand Pollination Effects in the Cherimoya (Annona cherímola).- Calif.Avoc.Soc. Yearb. 1941:94-98.
- Purificación Rosell Garcia, Victor Galán Saúco, Pedro M. Hernández Delgado 2007 . Cultivo Del Chirimoyo En Canarias. Cuadernos de divulgación. Departamento de Fruticultura Tropical Instituto Canario de Investigaciones Agradas. ICIA. Gobierno de canarias consejería de agricultura, ganadería, pesca y alimentación. Espana..
- FINCA EXPERIMENTAL "LA NACLA", 2003. Introducción al cultivo del chirimoyo
Enlaces externos
Tracheobionta
De Wikipedia, la enciclopedia libre
Tracheobionta es el nombre del
taxón de plantas que abarca a las
traqueófitas, también llamadas
plantas vasculares. Son organismos formados por
células vegetales, que poseen un ciclo de vida en el que se alternan las generaciones
gametofítica y
esporofítica, siendo esta última la fase dominante (sobre quien actúa más presión de selección natural); cuya fase esporofítica es fotosintética e independiente, y tiene tejidos y sistemas de órganos; está organizada en un "
cormo" (sistema que posee vástago aéreo, raíz subterránea y un sistema de conducción vascular que los vincula) que es a lo que comúnmente se refiere la gente cuando dice "planta"; cuya fase gametofítica es reducida y puede ser desde un "
talo" (cuerpo no organizado en tejidos ni órganos) en helechos y afines, hasta unas pocas células protegidas y nutridas por el
esporófito, en gimnospermas y angiospermas. La selección natural dirigió fuertemente la evolución de las traqueófitas hacia una menor dependencia de las condiciones ambientales sobre la tierra para la reproducción y la dispersión, característica que entra en evidencia al comparar las traqueofitas más antiguas (Lycophyta) con las más modernas (plantas con flores).
A lo largo de la historia se ha designado a este grupo con distintos nombres, como
cormófitas o
cormofitas (nombre científico
Cormophyta o
Cormobionta),
embriófitas sifonógamas (nombre científico
Embryophyta sifonogama), etc. Su nombre más común,
Tracheophyta, proviene de
tracheo (en referencia a las traqueidas, especializadas para el transporte de líquidos dentro de la planta) y
phyta, raíz de origen griego que significa "planta". Se lo castellaniza como
traqueofitas o
traqueófitas.
La evidencia de los
análisis moleculares de ADN hoy en día demostró que las traqueófitas son un grupo
monofilético (que comprende a todos los descendientes de un ancestro común) dentro de las
embriófitas, mientras que las
briofitas son
parafiléticas con respecto a las traqueofitas. Esto quiere decir que probablemente las traqueófitas sean descendientes de plantas muy parecidas a los briófitos, con el gametofito siendo la fase dominante, y el esporofito sin ramificar y dependiente nutricionalmente del gametofito.
Dentro de las traqueófitas hay dos linajes principales,
Lycophyta y
Euphyllophyta, diferenciadas principalmente por la forma de construcción de sus hojas (en las licofitas son lycofilos y en las eufilofitas son eufilos, los eufilos se corresponden aproximadamente con los megafilos, aunque en algunos grupos pueden estar reducidos en forma secundaria a su adquisición). Las eufilofitas a su vez comprenden dos grandes linajes,
Monilophyta (helechos, equisetáceas y psilotáceas) y
Spermatophyta, que se diferencian entre sí porque el primero tiene gametofitos de vida libre y el segundo los tiene encerrados en la semilla y el grano de polen. A su vez las espermatofitas están formadas por dos grandes grupos monofiléticos vivientes,
Gymnospermae y
Angiospermae o Magnoliophyta, que se diferencian entre sí porque el primer linaje tiene las semillas a la vista sobre la hoja fértil, mientras que el segundo tiene las semillas encerradas dentro de las paredes de la hoja fértil o carpelo.
Aún se siguen estudiando a las
Lycophyta y
Monilophyta como agrupadas en el grupo parafilético de los "helechos y afines" o
Pteridophyta.
Las traqueofitas son un grupo importante tanto por dominar la mayor parte de los ecosistemas terrestres como por ser muy utilizadas por el hombre.
El esporofito de las plantas vasculares
El cuerpo vegetativo
El cuerpo vegetativo del
esporófito es un "
cormo" (vástago + raíz + sistema de conducción vascular, hecho gracias al engrosamiento de las paredes celulares de células alargadas), que además de los tejidos vasculares posee tejidos de protección y tejidos de sostén, y que crece gracias a la acción de sus meristemas
anatomy/11.html anatomy/14.html anatomy/13.html.
El "
cormo" está especializado para la vida terrestre. Consta de vástago, raíz y un sistema de conducción vascular que los vincula. El
vástago es la región del cormo que se especializa en la fotosíntesis, y que utiliza para ello el agua y las sales minerales que le llegan de la raíz. La
raíz es la región del cormo que se especializa en la absorción de agua y sales, y que utiliza para ello los azúcares proporcionados por el vástago. Esta especialización es posible gracias a que el cormo posee un
sistema de conducción vascular formado gracias a que las paredes de la célula vegetal pueden volverse ricas en lignina, compuesto que les otorga dureza.
[editar] Meristemas
El cormo crece gracias a la actividad de sus
meristemas (del griego
meros: dividir) que son un grupo de células en estado embrionario permanente, capaces de dividirse indefinidamente, formando tejidos que en su juventud son indiferenciados
anatomy/11.html anatomy/14.html anatomy/13.html .
Los principales tipos de meristemas son dos:
- en todos los cormos encontraremos en los ápices de los tallos y las raíces, meristemas primarios responsables del crecimiento primario del esporofito.
- en los cormos con crecimiento secundario del tallo, encontraremos meristemas secundarios a lo largo del tallo, encargados de su crecimiento en grosor.
[editar] Sistemas de tejidos
El botánico J. Sachs en el siglo XIX, distinguió tres sistemas principales de tejidos en el cormo del esporofito de las cormófitas, clasificados según su función en la planta: tejidos de protección, tejidos fundamentales, y tejidos vasculares.
[editar] Tejidos de protección
Los tejidos que cumplen la función de protección forman la capa más externa del cormo.
- Si el cormo sólo posee crecimiento primario, encontraremos una epidermis, cubierta por una capa de cutina (lípido complejo que evita la pérdida de agua en la vida terrestre pero también evita el intercambio gaseoso con el medio ambiente), y con estomas Allies/Psilophyta/Psilotum nudum/Stem xs/Epidermis stomata.html y lenticelas (ambos aseguran el intercambio gaseoso con el ambiente).
- En los taxones con crecimiento secundario del tallo, la epidermis con cutina es reemplazada durante el crecimiento secundario, por una peridermis parcialmente impermeabilizada con suberina (lípido muy parecido a la cutina, responsable de la formación del súber o corcho).
Los tejidos fundamentales forman un sistema continuo y están conformados principalmente por los diversos tipos de
parénquimas (del griego: "carne de las
vísceras"), a los que se asocian los tejidos de sostén (como colénquima y esclerénquima).
Los
tejidos de sostén, como su nombre lo indica, cumplen la función de mantener la estructura de la planta, función que cumplen gracias a la
lignina presente en la pared celular. Los tejidos de sostén más comunes son:
- Colénquima (del griego: goma, cola, nombre dado por la facilidad con que las paredes celulares se hinchan al hidratarse) formado por células vivas.
- Esclerénquima (del griego escleros: duro, nombre dado por sus gruesas paredes muy duras y resistentes) formado por células casi siempre muertas a su madurez.
[editar] Tejidos vasculares
Son los responsables del transporte de líquidos y sustancias por todo el cuerpo del vegetal, en el que se distinguen el
floema (del griego
floeos: "yema de la corteza, propio de la corteza", especializado en transporte de azúcares) y el
xilema (del griego
xylos: "lignificarse", formado por células muertas tubulares, abiertas o cerradas en sus extremos, de paredes muy lignificadas, que forman un
haz vascular especializado en transporte de agua y sales)
[1] [2] [3] [4] [5]. Estos tejidos son complejos, y a menudo, están asociados a otros (parenquimáticos y de sostén). Los tejidos vasculares se ubican dentro de los tejidos fundamentales de manera diversa según los distintos órganos de la planta (raíz, tallo, etcétera), que en un corte transversal forman patrones (llamados
estela), que tienen importancia sistemática. Así el tallo de la mayor parte de las monocotiledóneas presenta un patrón o estela llamado
atactostela, el tallo (primario) de las dicotiledóneas y de las coníferas en cambio, presenta una
eustela, y el de los helechos y afines presenta una gran diversidad que permite diferenciar a las familias entre sí.
La reproducción en las plantas vasculares
 Diagrama esquemático del ciclo de vida de las plantas vasculares (Tracheobionta).
Referencias:
n : generación haploide,
2n : generación diploide,
m! : mitosis,
M! : meiosis,
F! : fecundación |
El esporofito adulto desarrolla los llamados
esporangios (estructuras multicelulares del esporofito dentro de las cuales se forman las esporas), donde se producirá la
meiosis que dará
esporas haploides. Las esporas al dividirse por mitosis se convierten en "gametofitos" (que serán un talo o sólo unas pocas células) que darán por meiosis las gametas masculina y femenina, la gameta femenina siempre será inmóvil y protegida al menos por el gametofito femenino dando en conjunto el óvulo, la gameta masculina será móvil o será transportada por agentes externos hasta el óvulo donde ocurre la fecundación, que dará un cigoto diploide. El nuevo esporofito se dividirá nutrido al principio al menos por el gametofito femenino, hasta dar el embrión formado por plúmula (donde está el meristema apical) y radícula (donde está el meristema radical), que al crecer forma el nuevo esporofito adulto recomenzando el ciclo. En los taxones sin hojas (
Psilotum) los esporangios se ubican directamente sobre el tallo, en los taxones con hojas, los esporangios se ubican en las hojas (convirtiéndose éstas en "
esporofilos").
Dentro de los esporangios se produce la meiosis que forma las
esporas haploides, que luego serán liberadas o no, y al dividirse por mitosis darán
gametofitos haploides. En las cormófitas más antiguas (como los helechos y afines) la fase gametofítica todavía es de vida libre, pero en las más recientes se ha vuelto totalmente dependiente del cuerpo vegetativo del esporofito para su nutrición y protección. Las pteridofitas (helechos y afines) pueden ser
isospóricas (todas las esporas iguales, darán gametofitos hermafroditas) o
heterospóricas (con dos morfos de esporas,
megasporas que darán gametofito femenino y
microesporas que darán gametofito masculino); en cambio las espermatofitas (gimnospermas y angiospermas) son todas heterospóricas. Cuando el esporofito es heterospórico, el gametofito se desarrolla por completo dentro de la espora, sin ser de vida libre. Los gametofitos producirán las
gametas femeninas y masculinas (del griego
gametes: marido). La
gameta femenina inmóvil espera ser fecundada dentro de una estructura donde está protegida y será nutrida (fenómeno de oogamia), esa estructura proviene del gametofito (en pteridofitas) o del gametofito y del esporofito (en espermatofitas). Al recibir a la
gameta masculina se produce la fecundación que dará el
cigoto diploide de un nuevo esporofito (del griego
zigos: pareja, yugo). El cigoto se empezará a dividir pasando por una fase de
embrión antes de convertirse en adulto, el embrión se nutre de sus cubiertas protectoras. Sólo en las espermatofitas el embrión puede mantenerse en estado de latencia (en forma de "semilla") hasta que las condiciones sean adecuadas (y se produzca lo que se conoce como "germinación"), las pteridofitas, más primitivas, se desarrollan directamente hasta adultas sin latencia. En el embrión ya se distinguen los meristemas primarios: el
meristema apical que da origen al vástago que también da origen a las hojas si las hay
anatomy/15.html, y un
meristema radical que da origen a la raíz primaria
anatomy/19.html. La raíz primaria o raíz embrionaria puede desarrollarse o morir, ya que el vástago también puede emitir raíces que en este caso se llamarán raíces adventicias.
Sistemática
Las cormófitas pueden dividirse en dos grandes grupos según el
sistema Engler:
- Grupo parafilético Pteridophyta. La hipótesis más aceptada hasta hace unos años sostenía que las pteridofitas son un grupo monofilético derivado de algún antecesor del alga verde. Hoy en día se sabe compuesta por dos líneas filogenéticas, son las más antiguas de las cormófitas actuales, comprende a los helechos y afines. Se caracterizan por poseer una alternancia de generaciones bien manifiesta. Las pteridofitas actuales están concentradas en los trópicos y áreas montanas húmedas. Está compuesta por las divisiones monofiléticas:
- División Lycopodiophyta
- División Monilophyta
- Superdivisión Spermatophyta. Las espermatofitas o antofitas pueden definirse como cormófitas donde la alternancia de generaciones se da en forma enmascarada, ya que el gametofito se desarrolla dentro de las estructuras del esporofito (incluso en los grupos más primitivos todavía pueden observarse anteridios y arquegonios). Comprende a las gimnospermas y angiospermas, en las siguientes líneas monofiléticas:
- División Gymnospermae
- División Magnoliophyta (angiospermas)
Los últimos análisis genéticos determinaron que la fiolegnia de las traqueófitas es la siguiente:
 Figura 5. Árbol filogenético actualizado de los diferentes grupos de pteridofitos y su relación con los demás grupos de plantas terrestres. |
En particular es de destacar que una evidencia molecular muy fuerte de la división basal entre licofitas y eufilofitas es la presencia en las eufilofitas de una inversión de unas 30 kilobases en el ADN del cloroplasto (Raubeson y Jansen 1992), lo notable es que inversiones de tantas kilobases son muy raras, y es raro que den productos viables, por lo que se convierte en evidencia muy fuerte de monofilia.
Evolución de traqueófitas
 Reconstrucción del esporofito de Rhynia. Aspecto general, corte longitudinal del esporangio, tétrada de esporas y detalle de espora con marca trilete. Nótense la ramificación dicotómica del tallo, la falta de hojas y raíces, y los esporangios terminales. |
Mucho de lo que sabemos acerca de las primeras plantas vasculares y su transición a la vida terrestre se encuentra en el registro fósil, ya que esos valiosos linajes están todos extintos.
Las traqueófitas ancestrales derivaron de las primeras
embriófitas, las primeras plantas que colonizaron la superficie terrestre. Todo indica que las primeras embriófitas eran pequeñas y muy simples en su estructura. En el caso del linaje que derivó en las plantas vasculares, el esporofito era básicamente un tallo de ramificación dicotómica, al principio de la altura de un fósforo, con el esporangio (donde ocurre la meiosis que da esporas haploides) producido en las puntas de las ramas. Estas plantas no tenían ni hojas ni raíces, en algunos casos (como
Rhynia del "
Rhynia Chert" en Escocia) la preservación de estas plantas es espectacular, y es posible discernir muchos detalles anatómicos, incluyendo estomas, esporas, y el tejido vascular dentro del tallo.
En los análisis basados en dichos fósiles, fue recientemente descubierto que los primeros
polysporangiofitos (plantas con esporofitos ramificados) en realidad no producían verdaderas células conductoras de agua (traqueidas) en el xilema, y por lo tanto deben haber dependido por completo de la presión de
turgencia para mantenerse erguidos. Las células verdaderamente conductoras de agua evolucionaron más tarde y caracterizan al clado de las traqueófitas o "verdaderas plantas vasculares" (Kenrick y Crane 1997a,
[1] 1997b
[2] ).
Las traqueidas son células elongadas con paredes engrosadas, muertas a la madurez. Donde una traqueida se conecta con la siguiente se observan unas aberturas características en la pared secundaria ("pits" en inglés) pero la pared celular primaria de las células que formaron las traqueidas se mantiene intacta, dando la "pit membrane". El agua por lo tanto debe atravesar las paredes primarias de las células para poder avanzar. En las primeras traqueófitas (representadas por
Rhynia)las traqueidas era de un tipo distintivo, con cierta "resistencia al desmoronamiento" ("decay resistance"), conferida por la lignificación de las fibras de celulosa, que estaba presente sólo como una capa muy delgada. Las paredes celulares más resistentes son las que caracterizan al clado de las
eutracheofitas, que include a todas las plantas vasculares vivientes (Kenrick y Crane 1997a
[1] ). En estas especies, las traqueidas fuertemente lignificadas permiten una conducción de agua más eficiente, y proveen resistencia interna, permitiendo a las plantas crecer mucho más altas.
Para leer una discusión acerca de la evolución de la predominancia del esporofito en detrimento del gametofito, ver
Evolución de embriófitas.
Las relaciones filogenéticas entre las líneas de plantas vasculares vivientes, mostradas en la figura de la sección sistemática, muestran una división basal, que ocurrió en el
Devónico temprano a medio (hace unos 400 millones de años), que separa al linaje de las
lycophytas actuales del linaje de las
eufilofitas actuales. Esta división está marcada por una considerable variedad de características morfológicas. Una bastante notoria es la presencia de esperma multiflagelado en las eufilofitas en oposición al esperma biflagelado de las lycophytas y de los linajes desprendidos de grupos más antiguos (los
briófitos sensu lato). Las dos únicas excepciones son
Isoetes y
Phylloglossum, donde el esperma multiflagelado se originó independientemente.
Las primeras plantas con semilla aparecieron en el Devónico tardío, y eso llevó a la típica flora del
Mesozoico dominada por gimnospermas. Los fósiles más antiguos de angiospermas encontrados hasta la fecha datan aproximadamente de hace 140 millones de años (principios del
Cretácico), y el número de especies encontrado aumenta de forma llamativa con el tiempo geológico, en comparación a otras plantas no angiospermas (el llamado "abominable misterio" de Darwin). Para una discusión sobre la radiación de las angiospermas ver
Evolución de angiospermas.
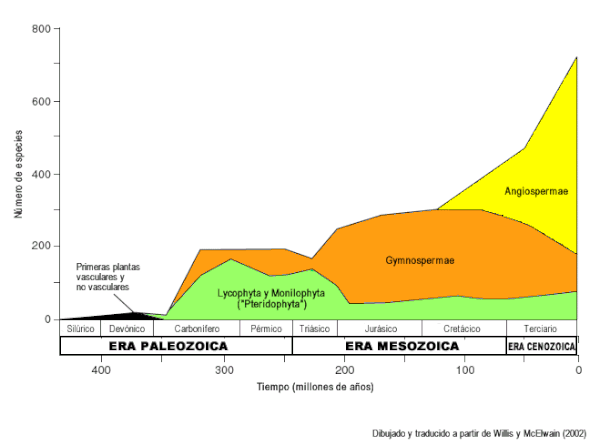 Registro fósil de aparición de los diferentes clados de plantas vasculares. Nótese la intensa radiación de las angiospermas en comparación con los helechos y gimnospermas. Dibujado y traducido a partir de Willis y McElwain (2002).[3] |
Importancia económica
Las traqueofitas son un grupo importante para el hombre, además de dominar la mayor parte de los ecosistemas terrestres: nos provee de medicinas, plantas ornamentales, fibras para papel y ropa, y la mayor parte de nuestros alimentos. Históricamente, la información sobre éstos y otros atributos de las traqueofitas fue esencial para el desarrollo de la civilización humana. La supervivencia dependía de conocer cuáles eran las plantas que eran buenas para comer, cuáles eran venenosas para la gente o para los animales que eran potencial alimento, cuáles eran buenas para armas y herramientas, cuáles podían curar, y cuáles podían ser útiles en muchos otros sentidos.
Enlaces externos
Bibliografía
- Judd, W. S.; Campbell, C. S.; Kellogg, E. A.; Stevens, P. F.; Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition. Sinauer Axxoc, USA.
- H. C. Bold, C.J. Alexopoulos y Th. Delevoryas (1989) Morfología de las Plantas y los Hongos. Omega, Barcelona, 911 pp., ISBN 84-282-0754-2
- Juan J. Valla, BOTÁNICA, Morfología de las Plantas Superiores, editorial Hemisferio Sur, Buenos Aires, Argentina, 1979.
- 2000. The PLANTS Database, database (version 5.1.1). National Plant Data Center, NRCS, USDA. Baton Rouge, LA 70874-4490 USA. http://plants.usda.gov
- Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (resumen en inglés aquí).
Referencias citadas
- ↑ a b Kenrick, P y PR Crane. 1997a. The origin and early diversification of land plants: A cladistic study. Smithsonian Institution Press, Washington, DC.
- ↑ Kenrick, P y PR Crane. 1997b. The origin and early evolution of plants on land. Nature 389: 33-39.
- ↑ Willis y McElwain. 2002. "Flowering Plant Origins". en: The Evolution of Plants, capítulo 6.
Lycophyta es una División de las
plantas vasculares que se caracteriza por poseer una alternancia de generaciones bien manifiesta (con esporofito y gametofito de vida libre), con el esporofito formando un cormo (vástago, raíz y un sistema de haces vasculares que los vincula) de tipo primitivo: el vástago consta de
tallo, y en las lycopsida también de
hojas de tipo
microfilo (de origen diferente de los megafilos de las euphyllophyta, probablemente derivadas de modificaciones de esporangios laterales, con un único
haz vascular), las raíces adventicias son originadas del vástago, las raíces tienen una característica ramificación dicotómica. En el corte transversal del tallo y la raíz los haces vasculares forman un patrón ("
estela") que al menos en las Lycopsida, en la raíz es una actinostela y en el tallo es derivada de la protostela con el xilema exarco, los esporangios son de inserción lateral, y en las lycopsida crecen en estrecha asociación con los microfilos estando ubicados en su cara adaxial, y usualmente con forma acorazonada.
Son las plantas vasculares más antiguas que existen en la actualidad, formando un clado (grupo monofilético) diferente de las
euphyllophyta en donde se reúne el resto de las plantas vasculares. Si bien su diversificación parece haber comenzado hace 200 millones de años aproximadamente, la diversificación que generó las especies conocidas en la actualidad parece haber empezado recién hace unos 80 millones de años (al terminar el Cretácico). En la actualidad todavía existe la clase Lycopsida (el resto de las clases ya extintas), siendo sus géneros más sobresalientes
Lycopodium,
Selaginella e
Isoetes.
Durante mucho tiempo se las ubicó junto con las
Monilophyta o helechos, formando la división
Pteridophyta, que hoy se sabe que es parafilética. Hasta hace poco tiempo se creía que las lycophytas eran tan antiguas como
Psilotum y
Equisetum, pero hoy se sabe que esos dos géneros son helechos que perdieron sus megafilos y otros caracteres de forma secundaria (después de haberlos adquirido).
Evolución
Las Lycophytas fueron especialmente diversas y abundantes en el
Carbonífero, y los restos de esas plantas representan nuestros mayores depósitos de carbón. En particular, ellas dominaron los pantanos costeros de las áreas bajas tropicales (DiMichele y Skog 1992, Bateman
et al. 1998).
Algunas lycophytas se volvieron árboles grandes, con crecimiento secundario que les permitía un aumento en grosor del tronco. Los tallos de estas plantas estaban cubiertos por hojas, lo que dejó las bases foliares características encontradas en los representantes fósiles. Estas plantas también desarrollaron los llamados "sistemas de raíces estigmarios" ("stigmarian root systems"), que se presume que fueron derivados de
rizomas en ese caso las raicillas dispuestas en espiral pueden ser hojas modificadas. Los patrones de crecimiento de estas enormes plantas aún son pobremente comprendidos (Bateman 1994, Bateman
et al. 1998), pero pueden haber crecido muy lentamente en altura al principio (mientras el sistema radical se establecía) y rápidamente más tarde. Pueden haber muerto luego de producir simultáneamente estróbilos en las puntas de todas sus ramas.
Diversidad
Hoy en día hay unas 1.200 especies de lycophytas, que pertenecen a varios linajes diferentes. De entre ellos, las especies rizomatosas de
Huperzia,
Lycopodiella y
Lycopodium son comúnmente encontradas en los bosques del
Hemisferio Norte. Estas plantas y sus parientes tropicales son homospóricas, lo que quiere decir que producen sólo un tipo de espora, que genera un
gametofito bisexual productor de
anterozoides y óvulos.
Las demás lycophytas vivientes (
Selaginella,
Isoetes) son heterospóricas, quiere decir que producen microsporas que generan gametofitos masculinos, y megasporas que generan gametofitos femeninos. Los
taxones heterospóricos forman un clado, que también tiene como diferencia característica una solapa de tejido de morfología foliar, llamada
lígula, asociada al lado adaxial de la base foliar.
Selaginella, con unas 700 especies, es más diversa en los trópicos, donde muchas especies crecen como epífitas.
Isoetes, con unas 150 especies, es el único resto viviente de un clado que incluía lycopodios gigantes del
Carbonífero, aunque probablemente derivó de plantas de este linaje que nunca alcanzaron el tamaño de
Lepidodendron y otros.
Isoetes ha retenido el cámbium y algo del crecimiento secundario, y tiene raicillas (rootlets) parecidas a las de los árboles extintos.
Referencias
- Pryer, Kathleen M., Harald Schneider, Alan R. Smith, Raymond Cranfill, Paul G. Wolf, Jeffrey S. Hunt y Sedonia D. Sipes. 2001. "Horsetails and ferns are a monophyletic group and the closest living relatives to seed plants". Nature 409: 618-622 (resumen en inglés aquí).
- Judd, W. S. Campbell, C. S. Kellogg, E. A. Stevens, P.F. Donoghue, M. J. 2002. Plant systematics: a phylogenetic approach, Second Edition.Sinauer Axxoc, USA.
- P. F. Stevens, 2001 en adelante, Angiosperm Phylogeny Website. Versión 9, junio del 2008, y actualizado desde entonces. http://www.mobot.org/MOBOT/research/APweb/


 Wikimedia Commons alberga contenido multimedia sobre Mangifera indica. Commons
Wikimedia Commons alberga contenido multimedia sobre Mangifera indica. Commons









